A propos du mannitol
Un certain nombre de patients atteints de la maladie de Parkinson prennent du mannitol en dehors de toute prescription médicale. Ces patients semblent vraiment bénéficier de l’ingestion de mannitol. Pourtant il y a des inconvénients sérieux comme les diarrhées intenses à doses excessives et tant que la flore intestinale ne s’est pas adaptée.
Il y a eu une étude de phase II sur l’impact du mannitol sur la maladie de Parkinson, la publication à ce sujet indique que l’étude n’a pas été satisfaisante mais l’échantillon était très petit et donc non significatif. Peut-être que le mannitol bénéficie seulement à un sous-ensemble de patients.
Il est intéressant d’examiner pourquoi le mannitol ou un médicament qui en serait dérivé, ne fait pas l’objet d’une recherche intense depuis que des études pré-cliniques ont montré son intérêt pour lutter contre la maladie de Parkinson. Les malades de la SLA trouveront des parallèles avec le TUDCA. C’est ce qu’on appelle en sociologie, un exemple de science (en tant que système de production de connaissances) inachevée.
Frustration face à une science inachevée
Le mannitol est une substance naturelle en poudre utilisée dans l’industrie alimentaire comme édulcorant. Les hôpitaux utilisent du mannitol par voie intraveineuse pour réduire la pression intracrânienne.
Entre 2012 et 2014, plusieurs études précliniques menées par une équipe de chercheurs de l'Université de Tel-Aviv, dirigée par les professeurs Dan Segal et Ehud Gazit, ont indiqué le potentiel thérapeutique du mannitol contre les amas de protéine alpha-synucléine.
Les chercheurs recherchaient une substance capable de décomposer ces amas de protéines in vitro, puis in-vivo sur les mouches des fruits (drosophiles). Ensuite, ils ont étudié des souris modèles de la maladie, là aussi, l'injection de mannitol a nettement réduit la formation d’amas de protéine alpha-synucléine . Après quelques semaines, les souris recevant du mannitol ont recommencé à courir et à grimper dans leurs cages. Suite à ces résultats encourageants, la prochaine étape évidente était l’expérimentation humaine… mais là, les choses se sont compliquées.
Le mannitol et les produits dérivés ne peuvent être brevetés facilement, car on en connaît de nombreuses variations naturelles. De plus, la valeur marchande des médicaments contre la maladie de Parkinson est énorme et devrait connaître un taux de croissance annuel important à l’avenir. Tout cela a rendu le mannitol peu attrayant pour tous les acteurs qui désirent activement breveter les résultats de leur recherche. Cela comprend évidemment les sociétés pharmaceutiques mais davantage encore les instituts gouvernementaux et les universités. En effet il n’y a pas besoin de brevet pour faire du profit en vendant un produit, c’est simplement un moyen de lutter contre les concurrents. Pour les acteurs de la recherche institutionnelle, breveter est le seul moyen de rentabiliser leurs travaux.
Une publication scientifique ayant étudié l’aspect social de l’abandon du mannitol par la recherche institutionnelle et sa continuation par le groupe CliniCrowd, rapporte les propos suivants:enter link description here
Mes recherches se sont arrêtées, l'argent s'est épuisé et j'ai constaté, à mon grand regret, que les projets sont tout simplement au point mort et qu'il y a encore des questions. Ce n’est pas que le projet soit inefficace ; souvent, j'ai vu de bons projets qui ont été abandonnés pour des raisons financières. À mesure que l’on gravit les échelons, cela devient de plus en plus évident, et je l’ai découvert moi-même… dans une société pharmaceutique israélienne, mais aussi au sein d’entreprises internationales comptant des dizaines de milliers de travailleurs. Vous vous rendez compte que le bien-être du patient dans ces lieux devient secondaire : l’argent, les investisseurs passent avant tout…. Je ne suis pas contre le fait de gagner de l'argent, c'est bien de gagner de l'argent, à condition de protéger avant tout le bien-être du patient. [Mais] lorsque le bien du patient est négligé pour des raisons financières – je veux dire, nous avons un très bon projet, mais nous ne l'avons pas encore breveté, et [le nouveau projet] est un meilleur traitement, mais nous ne le divulguerons pas au monde jusqu’à ce que nous le brevetions, et nous renouvellerons le brevet [de l'ancien médicament], même si nous savons que nous avons déjà quelque chose de mieux. Et ma conscience a eu du mal à accepter ce comportement.
De nouvelles formes d'activisme
La conjonction de chercheurs frustrés, de malades très bien informés et l’existence de réseaux sociaux permet la naissance de structures qui iront plus loin que le simple activisme ayant pour but de « financer la recherche » puisque la recherche de toute façon ne se fera que sur des résultats brevetables.
Les traitements peuvent également être abandonnés, notamment quand un concurrent propose une version générique ou quand le processus de fabrication devient trop coûteux alors que la plupart des médicaments sont aujourd’hui déjà sous-traités en particulier en Inde.
Ainsi, les premiers militants du sida étaient motivés en partie par leur conviction que les acteurs traditionnels de la recherche ne se soucieraient pas d’une maladie frappant principalement les groupes marginalisés et stigmatisés (Indyk et Rier, 1993). De même, les militants français de la dystrophie musculaire, ont pensé à une époque qu’ils étaient abandonnés par le corps médical et les scientifiques (Callon et Rabeharisoa 2003, p. 197). Dans les deux cas, les militants ont conclu que leur sort était entre leurs mains.
Le problème principal auquel se heurtent ces groupes n’est pas de trouver un médicament plus efficace que les médicaments existants, les systèmes de criblage automatisés (high throughput assay) et aujourd’hui les analyses in-silico, permettent d’obtenir de bons candidats rapidement. Le problème principal est plutôt de démontrer aux autorités de régulation, aux médecins et aux patients l’efficacité de ce médicament chez l’homme. C’est là que se situe l’essentiel des coûts d’introduction d’un nouveau médicament.
Quand un militaire s’empare du problème
Insatisfait de ses options de traitement, un patient et ancien général Israélien, Dan Vesely, a demandé a regroupé autour de lui, un petit groupe d’entrepreneurs et de chercheurs pour rechercher les recherches publiées sur la maladie de Parkinson. Ils ont vite remarqué l’étude intéressante de 2013, publiée et oubliée, sur l'effet possible du mannitol sur les patients atteints de la maladie de Parkinson.
Dan Vesely et certains partenaires ont contacté le professeur Dan Segal de l'Université de Tel-Aviv, qui avait codirigé l'équipe de recherche, et ont demandé à se rencontrer. Vesely raconte que le professeur Segal déclare au cours de la rencontre que tout sa recherche avait simplement été mis de côté, car il n’y avait aucune incitation financière pour les sociétés pharmaceutiques.
Dan Vesely veut alors de tester le mannitol sur lui-même, mais ses partenaires l’en dissuadent. Leur argument est que cela serait inefficace, et qu’il vaudrait mieux organiser un ensemble d’essais cliniques classiques. Au lieu de cela, ils ont accepté de tester le mannitol sur un certain nombre de patients atteints de la maladie de Parkinson.
Faute d’une société pharmaceutique disposée à investir dans la recherche clinique, ils cherchent alors à lancer leur propre société : CliniCrowd.
Inspiré par des projets de crowdsourcing similaires comme PatientsLikeMe, ou d’autres plus informels, le groupe créé un site Web destiné aux patients atteints de la maladie de Parkinson qui acceptent de prendre du mannitol régulièrement pendant une période prolongée. La société demande aux patients s'inscrivant sur le site de répondre à une enquête mensuelle détaillée sur leur santé, les symptômes de la maladie de Parkinson et la façon dont ils prenaient du mannitol.
Cette alternative ne constitue pas un véritable substitut aux essais cliniques « appropriés ». La plateforme d'enquête ne dispose pas d'un groupe témoin et la surveillance des patients est effectuée via le site de manière volontaire et indépendante, et non par un médecin. Néanmoins, ils génèrent des données préliminaires permettant de justifier la nécessité d’une recherche clinique plus formelle. Les fondateurs de CliniCrowd ont initialement envisagé de commercialiser directement le mannitol, mais ont décidé de ne pas le faire, pour éviter des conflits avec leurs recherches.
En fait, même si les perspectives de rentabilité de CliniCrowd sont relativement limitées, ils ont néanmoins choisi de l’enregistrer en tant que société plutôt qu’en tant qu’organisation à but non lucratif. En effet, les organisations à but non lucratif israéliennes sont strictement réglementées notamment dans le processus d’obtention d’un financement.
Ils ont créé la société en août 2016 et en début 2021, 2 480 patients s'étaient inscrits sur la plateforme dédiée à la recherche sur le mannitol pour la maladie de Parkinson. Parmi eux, 1 364 (55 %) avaient rempli des questionnaires à plusieurs reprises. Il était également possible de partager les données stockées avec le médecin traitant.
La principale valeur de cette action consistait à attirer des scientifiques accrédités pour mener des essais à plus grande échelle. Les données initiales de CliniCrowd ont suscité la pression du public, conduisant à leur tour à une étude clinique formelle, lancée en 2018 au centre médical Hadassah de Jérusalem. Cette étude (https://clinicaltrials.gov/ct2/show/NCT03823638), menée grâce à un financement public, examine les effets du mannitol sur la maladie de Parkinson. Même si l’étude de phase II ne montre pas, pour des raisons statistiques, une amélioration des patients recevant du mannitol par rapport au groupe de contrôle, c’est un premier effort. Après tout si les effets du mannitol sont manifestes à très court terme, il ne s’agit pas d’un essai en double aveugle.
Sans la pression exercée par le public, il est peu probable qu’un essai ait été lancé. Les résultats des travaux de CliniCrowd, n'ont pas remplacé la recherche clinique, mais contrairement à l'étude initiale sur le mannitol sur les mouches des fruits et les souris, ils ont créé un ensemble de données sur les patients difficiles à ignorer.
Evolution du discours de CliniCrowd
La position de CliniCrowd vis-à-vis de l’establishment biomédical a évolué au cours de sa courte histoire. Dans les premiers entretiens, les fondateurs ont vivement critiqué l’industrie pharmaceutique. Les premiers supports promotionnels de CliniCrowd, destinés au grand public, soulignaient à plusieurs reprises la réticence de l'industrie pharmaceutique à examiner l'effet du mannitol sur les patients atteints de la maladie de Parkinson. Lors d’une conférence de neurologues début 2017, les délégués de CliniCrowd n’ont eu que quelques minutes pour présenter, et la plupart des participants à la conférence ont ignoré leur discours.
Cependant, au fur et à mesure que le travail de terrain avançait, il y a eu un changement dans l’attitude des employés de CliniCrowd, dans le discours de l’entreprise et dans l’attitude des médias.
Le choix des termes « compléments alimentaires » et « aliments fonctionnels » plutôt que « médicament » reflétait la décision tactique de CliniCrowd de cesser de résister aux sociétés pharmaceutiques, en faveur de la création d'un canal d'influence et d'une action complémentaire, en recadrant le mannitol comme une nouvelle substance dans le marché des compléments alimentaires.
Cette tendance conforte l’analyse de Hess (2016, pp. 46-47) selon laquelle les challengers déplacent leur attention du remplacement des acteurs industriels vers la complémentarité. En cela, cela fait également écho à la recherche sur le sida, où des militants initialement profondément hostiles aux institutions scientifiques et pharmaceutiques ont finalement choisi de coopérer avec eux (Petersen et al. 2019 ; Harrington 1997 ; Maguire et al. 2004).
Présenter le mannitol en aliment fonctionnel s'est avéré une manœuvre payante. En effet, des chercheurs ont noté quelques années plus tard un changement d’attitude parmi les médecins « C'est un complément alimentaire. Cela n'aide peut-être pas, mais ce n'est pas nocif » .
Comme un médecin l'a expliqué :
Je pense qu'aucun médecin n'aime que le patient vienne et dise : « Écoutez, j'ai trouvé un traitement. » La plupart du temps, je dois m'assurer qu'il a les pieds sur terre et je dois lui expliquer pourquoi, très probablement, en son cas, ça ne marchera pas. C'était aussi ma première réponse au mannitol, une résistance totale, ne pas vouloir qu'ils le prennent... L'attitude change quand il y a déjà des informations et une masse de patients qui ont rassemblé et documenté leur utilisation de manière ordonnée. De plus, ils ne sont pas venus dire que c'était un médicament magique, mais plutôt qu'il pouvait aider à soulager certains symptômes... Je suggère aux patients, surtout au début, de se renseigner sur le mannitol. Je ne l’exclus certainement pas, bien au contraire.
les cas de science inachevée abondent.
Cependant il ne s’agit là que d’un exemple plutôt réussi, même si l’on se limite à la maladie de Parkinson, les cas de science inachevée abondent.
Il serait intéressant d’étudier les polyphénols tels l’Acide protocatéchique que l’on trouve dans divers produits naturels, la baicaléine, epigallocatechin-3-gallate. Les ingrédients actifs du ginseng, en particulier le Rb1, peuvent agir pour stabiliser les oligomères αSN non toxiques avec une teneur négligeable en feuilles β.
Des composés de pyrazole peuvent moduler les oligomères αSN. Cela conduit à l’efficacité confirmée de l’Anle138b dans différentes études in vitro et in vivo, qui corroborent également sa capacité à stopper la progression de la maladie et à prévenir la neurodégénérescence. Actuellement, Anle138b est en essai clinique de phase I.
Il est intéressant de noter que la β-synucléine, la protéine de 134 résidus de la famille des synucléines, a montré sa capacité intrinsèque à prévenir la fibrillation de l'αSN par liaison compétitive pour interférer avec les étapes de nucléation et d'agrégation.
Conclusion
CliniCrowd ou PatientsLikeMe ne sont pas forcément les seuls possibilitées d’activisme de malades, mais ce sont des exemples intéressants qui montrent qu’il ne suffit pas de « financer la recherche » il faut aller plus loin.
Discutez en sur le forum maladie de Parkinson
 The study shows systematic differences and potential directional biases between research and real-world datasets. Patients in research populations are diagnosed much earlier, start levodopa and other Parkinson's medications earlier, and show slower changes in clinical scales of motor and cognitive progression. Real-world-based populations are diagnosed at older ages, start medications later than research cohorts, and experience more rapid changes in clinical scales.
The study shows systematic differences and potential directional biases between research and real-world datasets. Patients in research populations are diagnosed much earlier, start levodopa and other Parkinson's medications earlier, and show slower changes in clinical scales of motor and cognitive progression. Real-world-based populations are diagnosed at older ages, start medications later than research cohorts, and experience more rapid changes in clinical scales.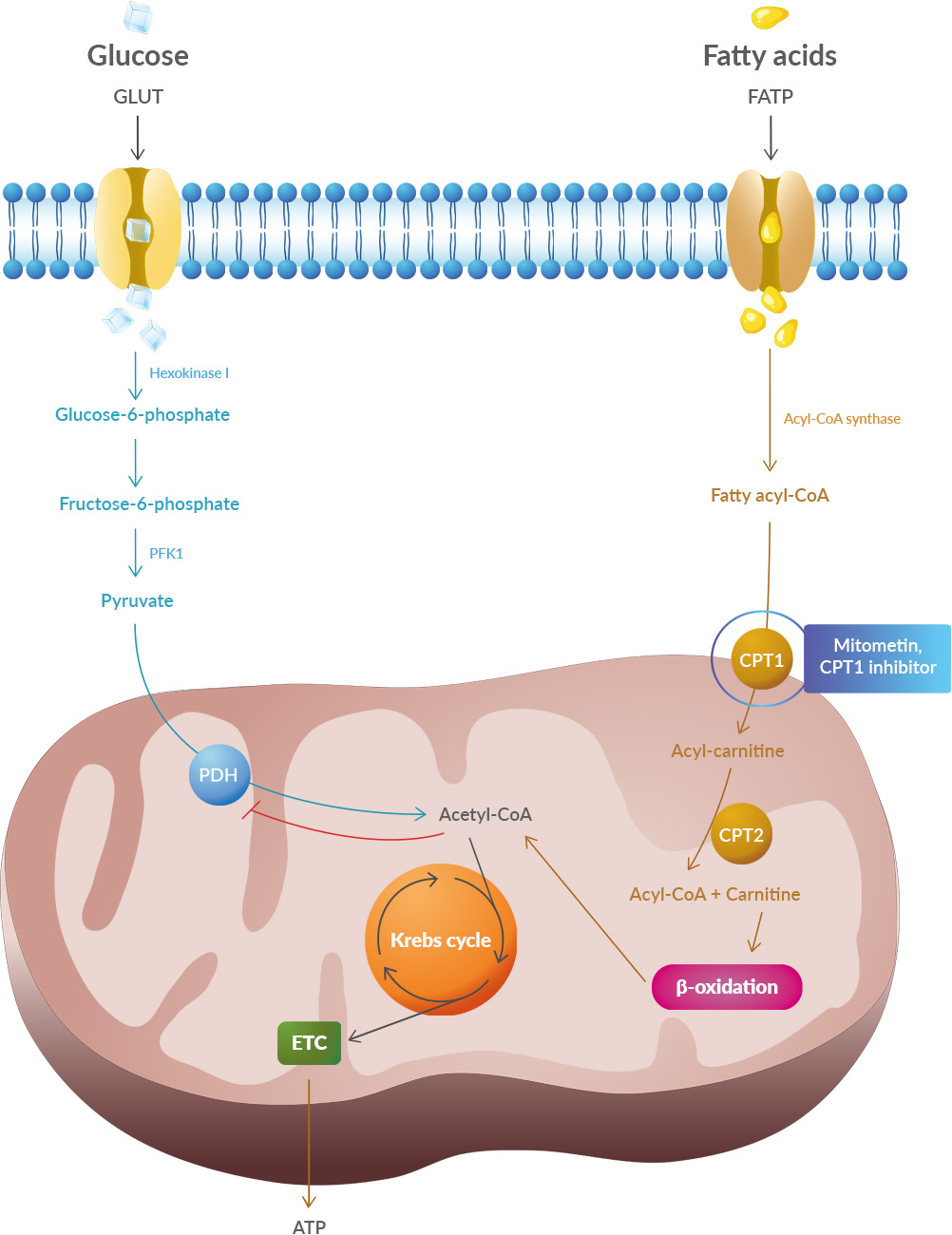 Studies have shown that oxidative stress and endoplasmic reticulum stress are correlated and can lead to protein misfolding (Abramov et al., 2020). Accumulation of misfolded proteins causes cellular damage and mitochondrial dysfunction and is associated with a range of neurodegenerative diseases, including ALS (misfolded SOD1, TDP-43, C9orf72) (McAlary et al., 2020), Parkinson's disease (misfolded α-synuclein) and Alzheimer disease (misfolded Aβ and Tau) (Abramov et al., 2020).
Studies have shown that oxidative stress and endoplasmic reticulum stress are correlated and can lead to protein misfolding (Abramov et al., 2020). Accumulation of misfolded proteins causes cellular damage and mitochondrial dysfunction and is associated with a range of neurodegenerative diseases, including ALS (misfolded SOD1, TDP-43, C9orf72) (McAlary et al., 2020), Parkinson's disease (misfolded α-synuclein) and Alzheimer disease (misfolded Aβ and Tau) (Abramov et al., 2020). It is also possible that there is an association of Parkinson's disease with a higher risk of important cardiovascular events like stroke and myocardial infarction (MI). 18F-dopamine is not the first tracer to demonstrate in identifying cardiac sympathetic dysfunction, however, Goldstein et al. are the first to demonstrate this relationship in a longitudinal, long-term prospective study of cardiac noradrenergic imaging in individuals with specific, self-reported nonmotor risk factors for Parkinson's disease.
It is also possible that there is an association of Parkinson's disease with a higher risk of important cardiovascular events like stroke and myocardial infarction (MI). 18F-dopamine is not the first tracer to demonstrate in identifying cardiac sympathetic dysfunction, however, Goldstein et al. are the first to demonstrate this relationship in a longitudinal, long-term prospective study of cardiac noradrenergic imaging in individuals with specific, self-reported nonmotor risk factors for Parkinson's disease. (Source: university of Toronto via Wikipedia)
(Source: university of Toronto via Wikipedia) Marc, comme de nombreux patients atteints de la maladie de Parkinson, était déjà équipé d'un dispositif de stimulation cervical profond (DBS). Aussi les chercheurs ont utilisé à la fois la stimulation cérébrale profonde, et la modulation de l’activité des motoneurones situés dans cette zone lombo-sacrée de la moelle épinière. Mais les chercheurs peuvent aussi traiter des patients non-équipé de ce dispositif. Dans les études sur les primates non humains, la neuroprothèse était reliée au cerveau des animaux pour provoquer une boucle de contre-réaction. Lorsque ces électrodes vertébrales étaient allumées, le gel de la démarche normalement observé chez Marc disparaisse presque, et ces effets positifs étaient observés que son simulateur DBS soit allumé ou éteint.
Marc, comme de nombreux patients atteints de la maladie de Parkinson, était déjà équipé d'un dispositif de stimulation cervical profond (DBS). Aussi les chercheurs ont utilisé à la fois la stimulation cérébrale profonde, et la modulation de l’activité des motoneurones situés dans cette zone lombo-sacrée de la moelle épinière. Mais les chercheurs peuvent aussi traiter des patients non-équipé de ce dispositif. Dans les études sur les primates non humains, la neuroprothèse était reliée au cerveau des animaux pour provoquer une boucle de contre-réaction. Lorsque ces électrodes vertébrales étaient allumées, le gel de la démarche normalement observé chez Marc disparaisse presque, et ces effets positifs étaient observés que son simulateur DBS soit allumé ou éteint.