Les oligonucléotides antisens (ASO) sont de petites séquences d'ADN capables de réduire l'expression d'un gène cible au niveau post-transcriptionnel, ce qui les rend intéressants pour neutraliser les produits génétiques mutants ou toxiques.
 Les progrès réalisés dans la chimie médicinale des ASO ont amélioré leur profil pharmacodynamique, permettant ainsi une administration sûre et efficace au système nerveux central. Les thérapies ASO pour la SLA se sont rapidement développées au cours des deux dernières décennies, et les ASO ciblant SOD1, C9orf72 et ATXN2 sont actuellement en essais cliniques pour les formes familiales ou sporadiques de SLA.
Les progrès réalisés dans la chimie médicinale des ASO ont amélioré leur profil pharmacodynamique, permettant ainsi une administration sûre et efficace au système nerveux central. Les thérapies ASO pour la SLA se sont rapidement développées au cours des deux dernières décennies, et les ASO ciblant SOD1, C9orf72 et ATXN2 sont actuellement en essais cliniques pour les formes familiales ou sporadiques de SLA.
L'injection directe dans le SNC permet à l'ASO de se distribuer dans tout le SNC, et les ASO ciblant SOD1 (Tofersen/Qalsody) ont démontré que l'administration intrathécale était une approche bien tolérée. De nouvelles approches d'administration, telles que la conjugaison des ASO à des nanoparticules lipidiques ou à des cholestérols, pourraient bientôt permettre une administration moins intrusive et pouvant être effectuée par davantage de professionels.
Les mutations du sarcome fusionné (FUS) sont à l'origine d'une forme rare et agressive de SLA, d'apparition précoce et souvent juvénile. FUS est une protéine de liaison à l'ARN essentielle à la réparation et au métabolisme de l'ADN, notamment à l'épissage et à la traduction de l'ARNm.
Alors que la plupart des formes de SLA présentent généralement une pathologie TDP-43, les tissus post-mortem de patients atteints de SLA présentent une agrégation intracytoplasmique en l'absence de pathologie TDP-43.
 Identifié par criblage in vitro, l'ASO ION363 développé par la société IONIS qui a aussi développé Tofersen, cible le 6e intron de FUS (SLA avec une mutation P525L).
ION363 réduit les taux de protéines de liaison à l'ARN insolubles et insolubles associées aux agrégats, telles que hnRNPA1 et ralentit la neurodégénérescence des motoneurones lombaires et la perte d'innervation de la jonction neuromusculaire.
Identifié par criblage in vitro, l'ASO ION363 développé par la société IONIS qui a aussi développé Tofersen, cible le 6e intron de FUS (SLA avec une mutation P525L).
ION363 réduit les taux de protéines de liaison à l'ARN insolubles et insolubles associées aux agrégats, telles que hnRNPA1 et ralentit la neurodégénérescence des motoneurones lombaires et la perte d'innervation de la jonction neuromusculaire.
L'inversion de la neurodégénérescence et la réduction de la prolifération chez les souris P525L ont motivé une demande d'IND (« usage compassionnel ») pour des tests chez l'homme porteur de mutations. Une demande d’IND a été approuvée par la FDA pour l’utilisation d’ION363 chez un patient atteint de SLA porteur d’une mutation P525L (âgé de 26 ans).
Jaci Hermstad, une jeune femme de 25 ans originaire de l'Iowa, a reçu un diagnostic de SLA-FUS, huit ans après avoir perdu sa sœur jumelle, atteinte de la même maladie, à l'âge de 17 ans. La famille Hermstad a contacté Project ALS et le Dr Shneider, qui étudiait le potentiel des ASO dans le traitement des patients atteints de SLA-FUS, pour savoir s'il existait des thérapies susceptibles d'aider Jaci.
« L'histoire des Hermstad a immédiatement attiré l'attention de nombreuses personnes talentueuses et bienveillantes, d'Ionis et du Dr Shneider, expert du gène FUS, aux experts réglementaires bénévoles, aux fabricants et aux conseillers universitaires », a déclaré Valerie Estess, directrice de recherche du Project ALS. Le courage de Jaci, et le travail d'équipe qu'elle a inspiré, peuvent désormais porter leurs fruits pour tous les patients atteints de SLA-FUS.
Grâce au financement de l'ALS Association et du Projet SLA, le Dr Shneider et son équipe de Columbia ont pu proposer le jacifusen à dix patients supplémentaires atteints de SLA-FUS au cours des deux dernières années, tout en suivant en parallèle les données de sécurité du médicament et les biomarqueurs pertinents pour la maladie.
Cela a conduit la Chambre des représentants des États-Unis à adopter le projet de loi de Jaci, autorisant les médecins à administrer l'ASO avant de réaliser des tests toxicologiques sur des rongeurs. Selon sa nécrologie, Hermstad a reçu 12 injections du médicament, appelé Jacifusen, entre juin 2019 et mars 2020 avant de décéder de la SLA le 1er mai 2020.
À l'autopsie, l'ION363 a été largement détecté dans les tissus du cerveau et de la moelle épinière, deux mois après la dernière perfusion. Les signes pathologiques de la SLA-FUS P525L ont diminué, notamment les inclusions cytoplasmiques neuronales positives, les agrégats insolubles de protéines de liaison à l'ARN et d'autres protéines, ainsi que la localisation nucléaire.
Un examen neuropathologique a été réalisé chez le premier participant, ainsi que chez un témoin non atteint de SLA et un patient atteint de SLA porteur de la mutation P525L n'ayant pas reçu de traitement. Par rapport au témoin atteint de SLA, le participant traité par ASO présentait moins de protéines totales et mutantes, y compris de protéines insolubles, dans la moelle épinière lombaire.
Shneider et son équipe à Columbia ont proposé du Jacifusen à dix patients supplémentaires atteints de SLA au cours des deux dernières années.
En juin 2021, Ionis a lancé un essai de phase 3 appelé ION, visant à traiter jusqu'à 77 patients dans le monde. Les participants sont âgés de 11 ans et plus, atteints de SLA causée par une mutation pathogène confirmée du gène ION363 et ne doivent pas être sous ventilation mécanique permanente au moment de l'inscription. Ils reçoivent des injections rachidiennes d'ION363 ou d'un placebo toutes les douze semaines, après une dose de charge à quatre semaines, pendant 61 semaines, suivies d'une prolongation en ouvert de 85 semaines. Le critère d'évaluation principal est l'évolution fonctionnelle selon l'échelle d'évaluation fonctionnelle de la SLA révisée et la durée de vie sans ventilation mécanique. Les critères d'évaluation secondaires incluent la qualité de vie, la fonction pulmonaire et musculaire, la survie et les modifications du biomarqueur des chaînes légères des neurofilaments. Réalisé à L'essai clinique, mené sur 24 sites en Amérique du Nord, en Europe, au Royaume-Uni, à Taïwan et en Corée, devrait se poursuivre jusqu'en juin 2026.
La deuxième partie de l'étude consiste en une période d'extension ouverte de 72 semaines au cours de laquelle tous les participants ont reçu du jacifusen.
Une caractéristique unique de cet essai est la mise en œuvre d'un « sauvetage ». Plus précisément, si un participant présente un déclin fonctionnel significatif au cours de la première partie, il sera transféré vers la deuxième partie/extension ouverte de l'étude. Cela semble suspect du point de vue statistique: L'étude ne conserve que les patients qui évoluent lentement!
Bien que la plupart des participants aient connu un déclin fonctionnel continu (mesuré par l’ALSFRS-R) après le début du traitement par jacifusen, l’un d’eux a présenté une récupération fonctionnelle objective sans précédent après 10 mois, et un autre est resté asymptomatique, avec une amélioration documentée des anomalies électromyographiques.
 The researchers specifically focused on molecules that could reduce or reverse stress granule formation, particularly those that act directly on stress granule proteins and may be useful as therapeutic agents. From the initial screening, lipoamide emerged as a novel, potent modulator of stress granules.
Once lipoamide was identified as a hit, the researchers sought to determine its effects in cells regarding specificity, potency, intracellular localization, and its effects on other cellular condensates.
The researchers specifically focused on molecules that could reduce or reverse stress granule formation, particularly those that act directly on stress granule proteins and may be useful as therapeutic agents. From the initial screening, lipoamide emerged as a novel, potent modulator of stress granules.
Once lipoamide was identified as a hit, the researchers sought to determine its effects in cells regarding specificity, potency, intracellular localization, and its effects on other cellular condensates.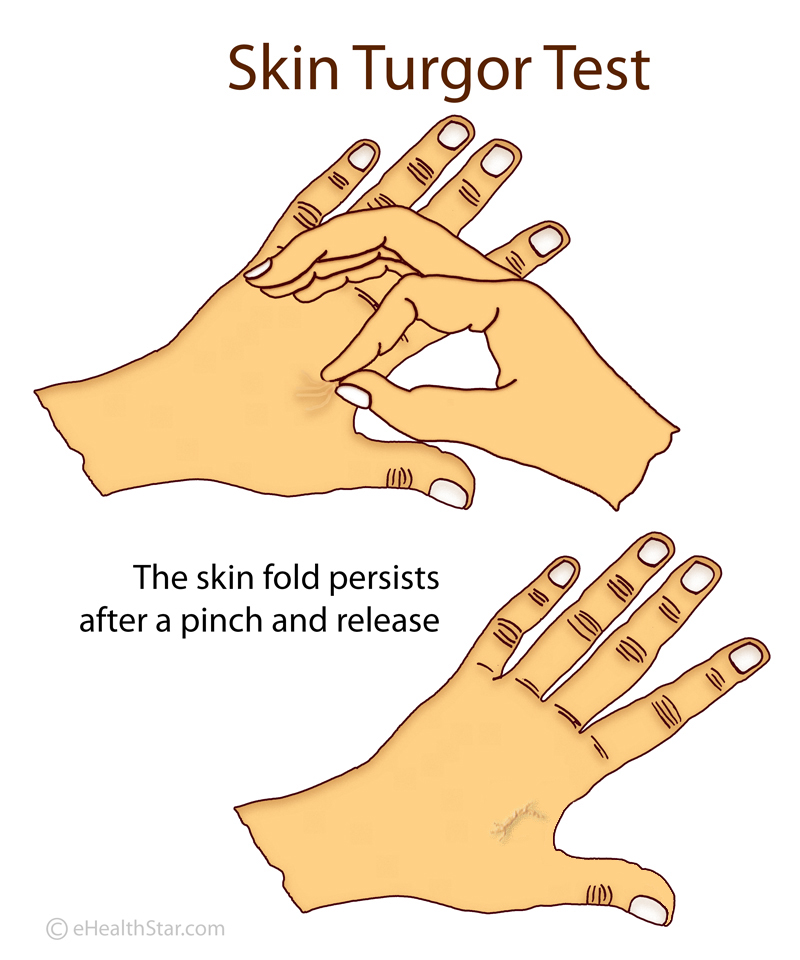 In healthy individuals, after a deformation or pinching, the skin quickly returns to its original shape. In patients with ALS, this return is slower. This is called the delayed return phenomenon (DRP).
In healthy individuals, after a deformation or pinching, the skin quickly returns to its original shape. In patients with ALS, this return is slower. This is called the delayed return phenomenon (DRP). Some people in the intervention group showed an astonishingly slower disease progression compared to the delayed-start group:
Some people in the intervention group showed an astonishingly slower disease progression compared to the delayed-start group: